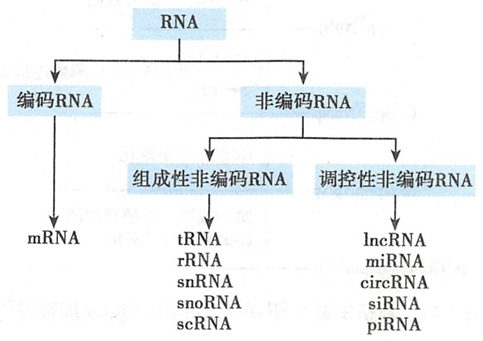
RNA
非编码RNA可以分为两类。
- 一类是确保实现基本生物学功能的RNA,它们的丰度基本恒定,故称为组成性非编码RNA(constitutive non-coding RNA)。包括
- 另一类是调控性非编码RNA(regulatory non-coding RNA),它们的丰度随外界环境(应激条件等)和细胞性状(成熟度、代谢活跃度、健康状态等)而发生改变,在基因表达过程中发挥重要的调控作用。
RNA通常以单链形式存在,较长的RNA可以通过链内的碱基互补配对形成局部的双螺旋二级结构以及复杂的高级结构。
一、mRNA
信使RNA(mRNA)。在生物体内,mRNA的丰度最小,仅占细胞RNA总重量的2%~5%。但是mRNA的种类最多。
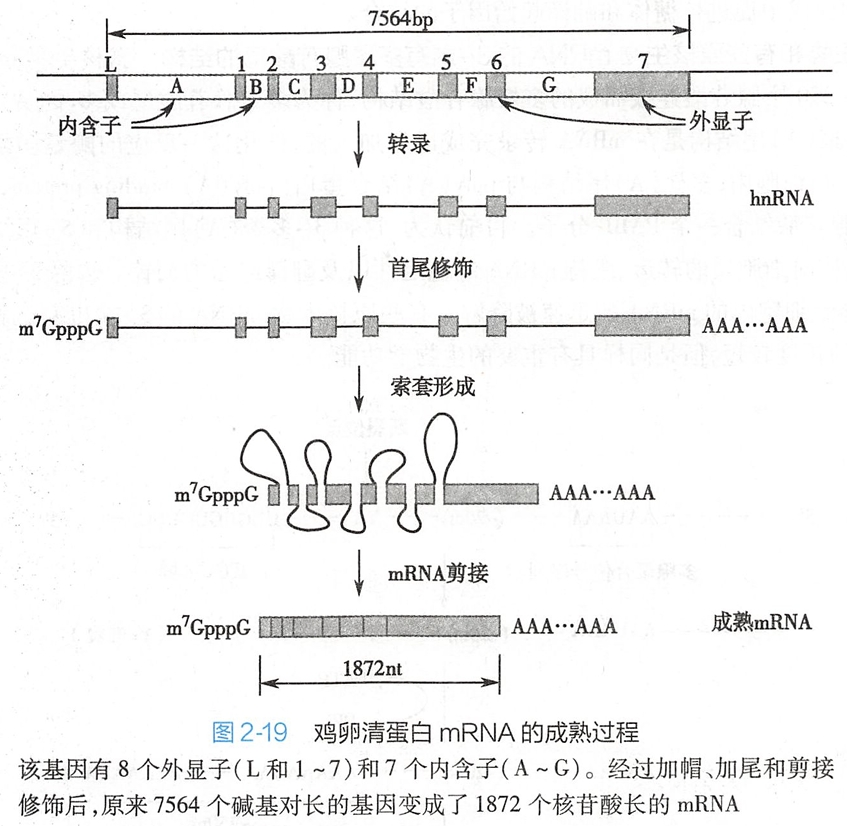
在真核细胞中,细胞核内新生成的mRNA初级产物被称为核不均一RNA(heterogeneous nuclear RNA, hnRNA)。hnRNA在细胞核内合成后,经过一系列的转录后修饰,剪接成为成熟mRNA,最后被转运到细胞质中。
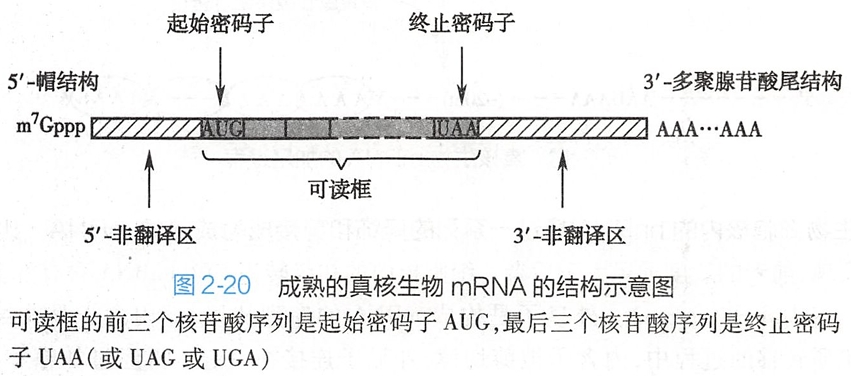
(一)真核细胞mRNA的5'-端有帽结构
大部分真核细胞mRNA的5'-端都有一个反式7-甲基鸟嘌呤-三磷酸核苷(m7Gppp)的起始结构,被称为5'-帽结构(5'-cap structure)(图2-17)。
5'-帽结构是鸟苷酸转移酶将鸟嘌呤三磷酸核苷(GTP)加到转录后的hnRNA的5'-端,形成了一个5'-5'三磷酸键,使mRNA的5'-端不再具有磷酸基团。(新生的hnRNA的长度达25~30bp时,就开始进行戴帽修饰。)
5'-帽结构下游的第一个和第二个核苷酸中C-2'的羟基通常也会被甲基化成为甲氧基戊糖,由此产生数种不同的帽结构(图2-17)。
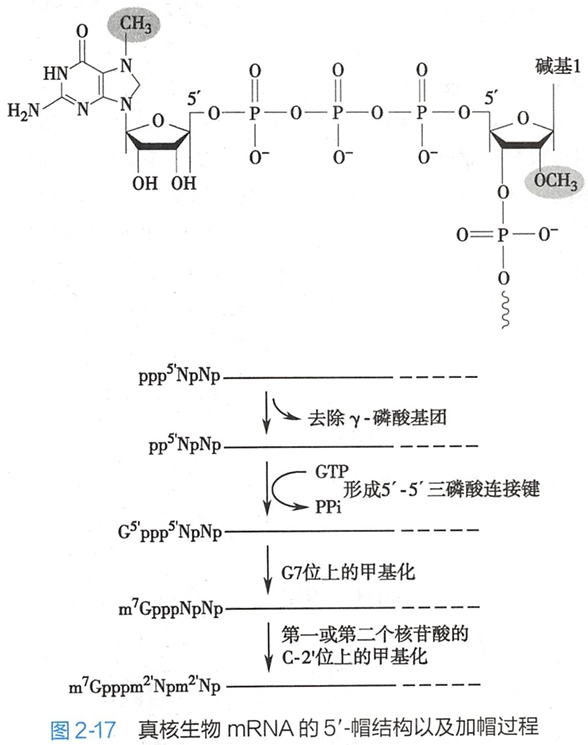
原核生物RNA没有这种特殊的 5'-帽结构。真核生物mRNA的5'-帽结构可以与一类称为帽结合蛋白(cap binding protein,CBP)的分子结合形成复合体。这种复合体有助于维持mRNA的稳定性,协同mRNA从细胞核向细胞质的转运,并促进核糖体与翻译起始因子的结合。
(二)真核细胞和有些原核细胞mRNA的3'-端有多聚腺苷酸尾
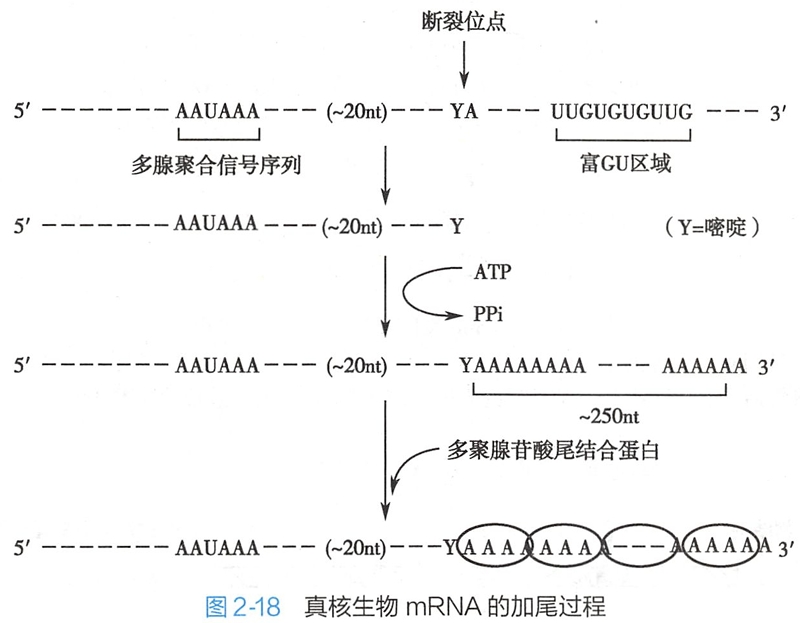
加尾(tailing)是指对hnRNA3'端的修饰过程 ,即在腺苷酸聚合酶的作用下,在3'端加上由200~250个腺苷酸组成的多聚腺苷酸(poly-A)的尾巴。poly-A与多个poly-A结合蛋白(PABP)结合。
加尾的作用:
- 一方面可使mRNA3'端稳定,防止被核酸酶水解,
- 另一方面有利于mRNA由核到细胞质的转运。
目前发现,决定mRNA前体分子加尾的信号序列存在于RNA3'端,即在发生多聚腺苷酸化位点的上游10~30bp处存在一高度保守的6核苷酸序列(在哺乳动物中为AAUAAA),在多聚腺苷酸化位点的下游常存在富含G和U的序列。这些序列可被含有腺苷酸聚合酶的复合物所识别,进行加尾。
(三)hnRNA
hnRNA先剪切内含子,再加帽加尾成mRNA。(我觉得,hnRNA合成时就加帽,再加尾,再剪切内含子成mRNA。) #?
二、tRNA
(一)稀有碱基
稀有碱基(rare base)是指除A、G、C和U外的一些碱基,包括双氢尿嘧啶(dihydrouracil,DHU)、假尿嘧啶核苷(pseudouridine,Ψ)和甲基化的嘌呤(m7G、m7A)等(图2- 21)。
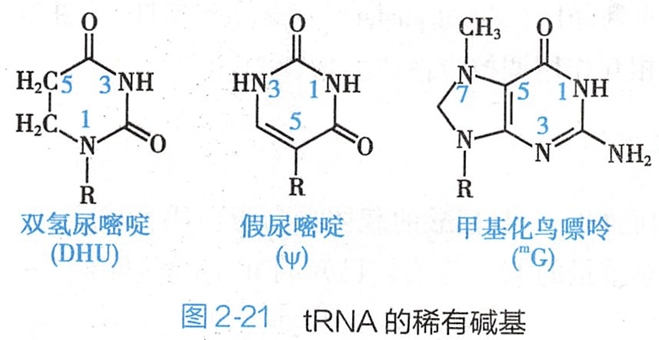
正常的嘧啶核苷是杂环的N-1原子与戊糖的C-1'原子连接形成糖苷键,而假尿嘧啶核苷则是杂环的C-5原子与戊糖的C-1'原子相连。tRNA中的稀有碱基占所有碱基的10%~20%。tRNA分子中的稀有碱基均是转录后修饰而成的。
(二)特定的空间结构
tRNA存在着一些核苷酸序列,能够通过碱基互补配对的原则,形成局部的链内的双螺旋结构。
在这些局部的双螺旋结构之间的核苷酸序列不能形成互补的碱基对则膨出形成环状或襻状结构。这样的结构称为茎环(stem-loop)结构或发夹(hairpin)结构。
由于这些茎环结构的存在,tRNA的二级结构呈现出酷似三叶草(cloverleaf)的形状(图2-22)。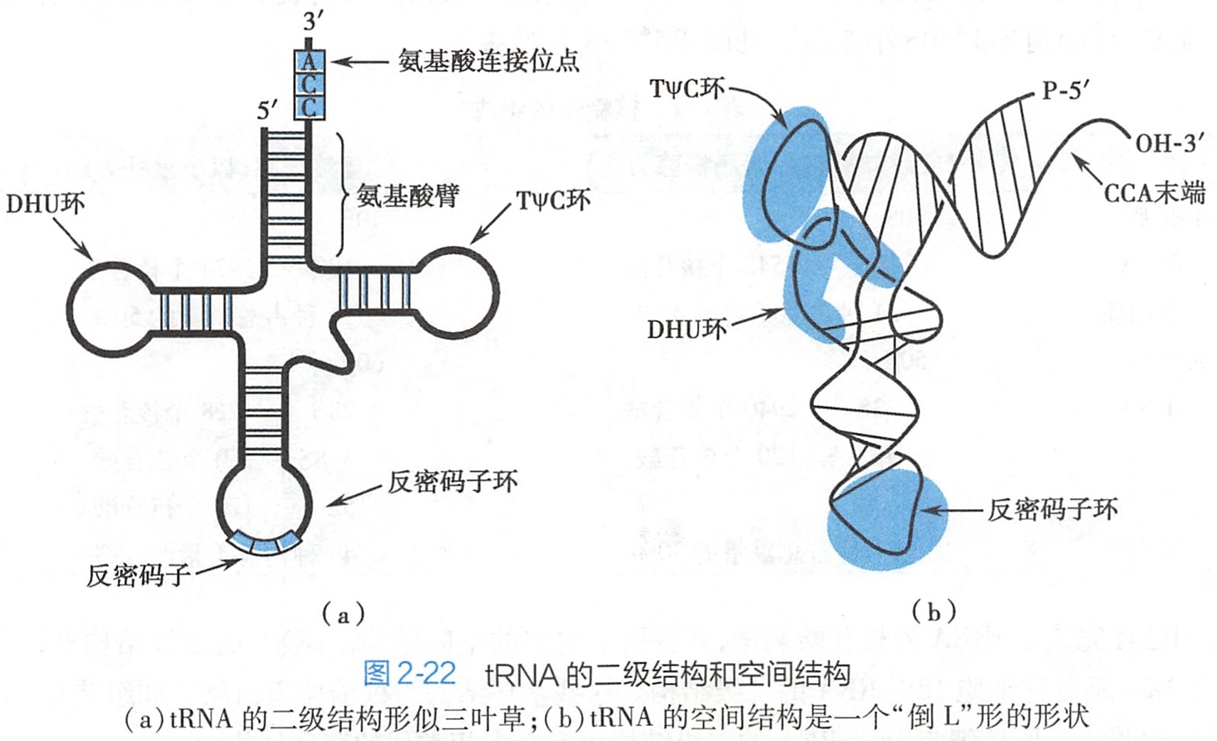
位于两侧的发夹结构含有稀有碱基,分别称为DHU环和TΨC环;位于上方的茎称为氨基酸臂(amino acid arm),亦称接纳茎;位于下方的发夹结构则称为反密码子环(anticodon loop)。
此外,在反密码子环与TΨC环之间还有一个可变臂。不同tRNA的可变臂的长短不一,从几个到十几个核苷酸数不等。除可变臂和DHU环外,其他部位的核苷酸数目和碱基对具有高度保守性。
所有的tRNA都具有相似的倒“L”形的空间结构。稳定tRNA的三级结构的力是某些碱基之间产生的特殊氢键和碱基堆积力。
(三)tRNA的3'-端连接着氨基酸
所有tRNA的3'-端都是以CCA三个核苷酸结束的,氨酰-tRNA合成酶将氨基酸通过酯键连接在腺嘌呤(应该是腺苷酸)A的C-3'原子上,生成了氨酰-tRNA,从而使tRNA成为了氨基酸的载体。
有的氨基酸只有一种tRNA,而有的氨基酸有几种tRNA作为载体,以适应mRNA上密码子简并性的需求。(这说明为了适应简并性的需求并没有只靠摆动性)
(四)反密码子
tRNA的反密码子环由7~9个核苷酸组成,居中的3个核苷酸通过碱基互补配对的关系识别mRNA上的密码子,因此被称为反密码子。
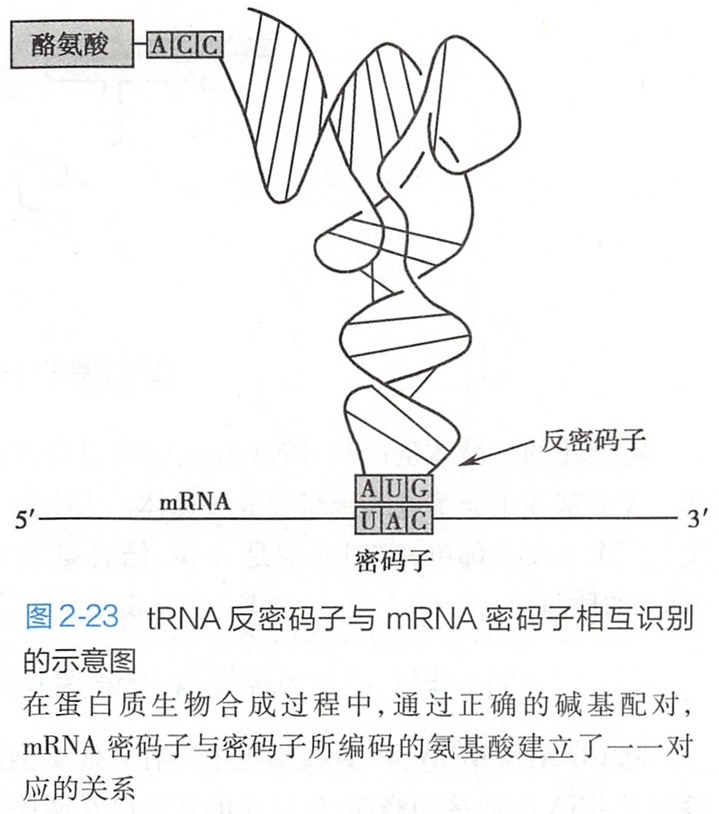
例如,携带酪氨酸的tRNA反密码子是-GUA-,可以与mRNA上编码酪氨酸的密码子- UAC-互补配对(图2-23)。
三、rRNA
核糖体RNA(ribosomal RNA,rRNA)是细胞中含量最多的RNA,约占RNA总重量的80%以上。

S是大分子物质在超速离心沉降中的沉降系数。
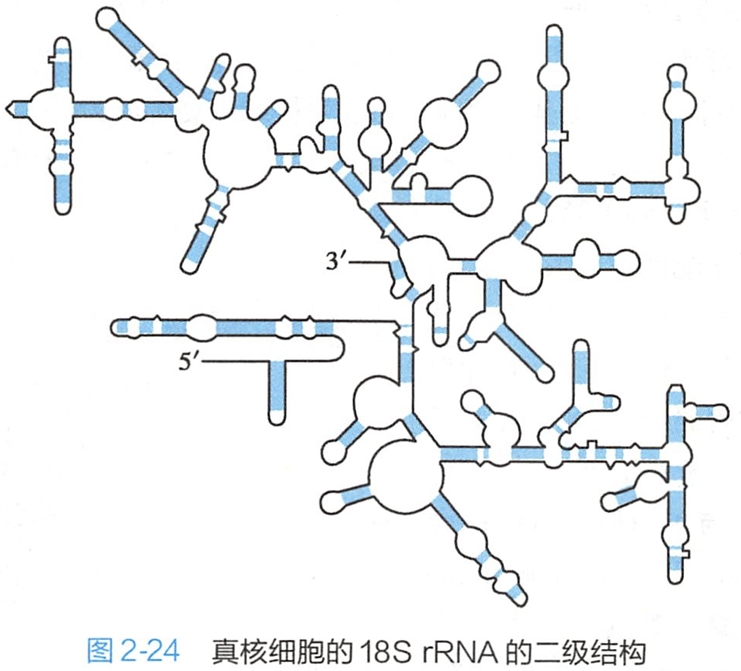
rRNA的二级结构有许多茎环结构,图2-24是真核细胞18S rRNA的二级结构。这些茎环结构为核糖体蛋白结合和组装在rRNA上提供了结构基础。原核细胞16S rRNA的二级结构也有众多相类似的茎环结构。
核糖体有三个重要的部位,它们分别是A位:结合氨酰-tRNA的氨酰位(aminoacyl site);P位:结合肽酰 tRNA的肽酰位(peptidyl site);E位:释放已经卸载了氨基酸的tRNA的排出位(exit site)(图2-25)。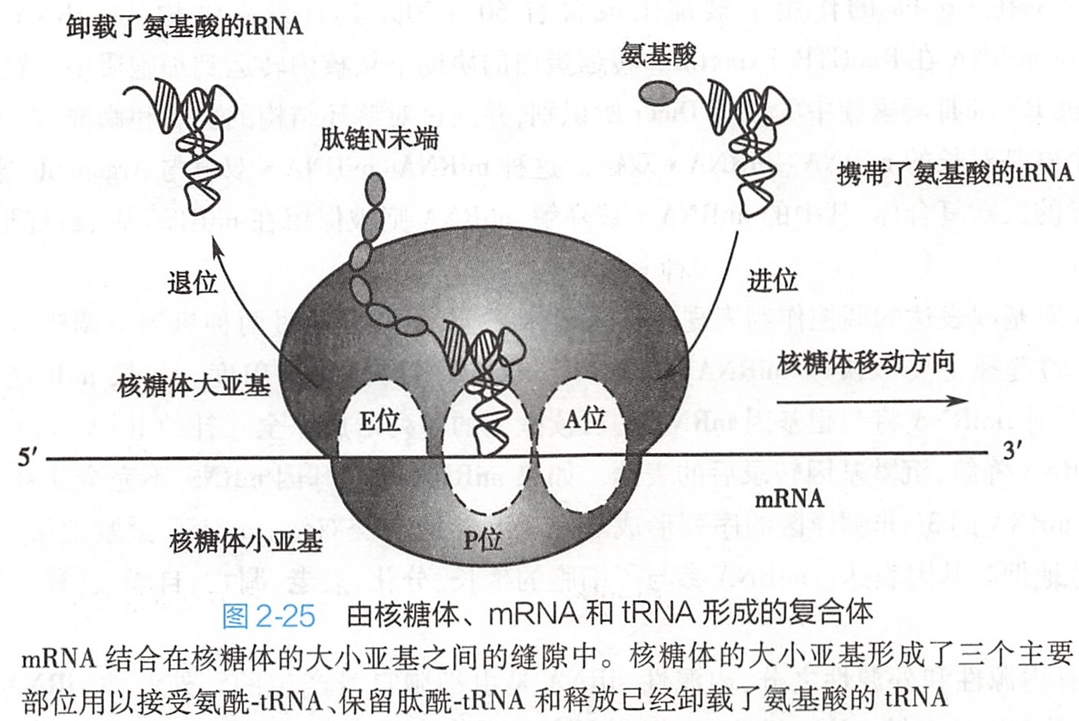
四、组成性非编码RNA
除tRNA和rRNA外,真核细胞中还有其他类型的组成性非编码RNA。组成性非编码RNA是保障遗传信息传递的关键因子。这些RNA作为关键因子参与了RNA的剪接和修饰、蛋白质的转运以及调控基因表达。
(一)催化小RNA
催化小RNA也称为核酶(ribozyme),是细胞内具有催化功能的一类小分子 RNA统称,具有催化特定RNA降解的活性,在RNA合成后的剪接修饰中具有重要作用(见第十四章)。
(二)核仁小RNA
核仁小RNA(small nucleolar RNA,snoRNA)定位于核仁,主要参与rRNA的加工。tRNA的核糖C-2'的甲基化过程和假尿嘧啶化修饰都需要snoRNA的参与。
(三)核小RNA
核小RNA(small nuclear RNA,snRNA)参与了真核细胞mRNA的成熟过程。一个snRNA与大约20种蛋白质组成了细胞的核小核糖核蛋白(small nuclear ribonucleoprotein, snRNP)。由于它们富含尿嘧啶,故命名为U-snRNA。研究比较清楚的snRNA有U1、U2、U4、U5、U6和U7。它们的作用是识别hnRNA上的外显子和内含子的接点,切除内含子。这些snRNA的5'-端有一个与mRNA相类似的5'-帽结构(见第十四章)。
(四)胞质小RNA
胞质小RNA(small cytoplasmic RNA,scRNA)存在细胞质中,与蛋白质结合形成复合体后发挥生物学功能。例如,SRP-RNA与六种蛋白质共同形成信号识别颗粒(signal recognition particle,SRP),引导含有信号肽的蛋白质进入内质网进行合成(见第十五章)。
五、调控性非编码RNA
调控性非编码RNA按其大小分为非编码小RNA(small non-coding RNA,sncRNA)、长非编码RNA(long non-coding RNA,IncRNA)和环状RNA(circular RNA,circRNA)。
虽然这一类RNA通常不编码蛋白质,但是它们仍然表现出了许多重要的生物学功能:转录调控、RNA剪切和修饰、RNA的翻译、蛋白质的稳定和转运、染色体的形成和结构稳定等,因此,在胚胎发育、组织分化、信号转导、器官形成等基本的生命活动中以及在疾病(如肿瘤、神经性疾病等)的发生和发展进程中都有非编码RNA的参与。
(一)sncRNA
通常认为,sncRNA的长度小于200nt。
sncRNA包括微RNA(microRNA,miRNA)、干扰小RNA(small interfering RNA,siRNA)和piRNA。
1.miRNA
微RNA是近年来研究较多的内源性sncRNA,它在真核生物中大量存在,长度在20~25nt之间。
在细胞核中,编码miRNA的基因由RNA聚合酶Ⅱ转录生成长度约为几千个碱基的初级转录本pri-miRNA。
在细胞核内,pi-miRNA在蛋白质复合体的作用下经过了第一次的加工。这个蛋白质复合体由Drosha和Pasha两个蛋白质组成,它们分别是RNase Ⅲ蛋白和双链RNA结合蛋白。pri-miRNA在Drosha的作用下被加工成含有60~70nt具有发夹结构的miRNA前体(pre miRNA)。
pre-miRNA在RanGTP/Exportin-5转运蛋白的协助下从核内转运到细胞质中。在细胞质中, pre-miRNA被RNaseⅢ酶家族中的成员Dicer所识别,并通过对茎环结构的剪切和修饰,在细胞质内形成大约20个碱基对长的miRNA:miRNA*双链。
这种miRNA:miRNA*双链与Argonaute家族蛋白形成RNA诱导的沉默复合体,其中的miRNA*被降解,miRNA则被保留在miRISC中,最终形成成熟的单链miRNA。
微RNA对基因表达的调控作用表现在转录后水平上,主要是通过两种机制下调靶基因的表达。这两种机制的选择主要取决于miRNA与靶基因mRNA序列的互补程度。
- 如果miRNA与靶基因mRNA完全互补,miRNA将与靶基因mRNA的可读框中的序列形成完全互补的RNA双链,miRISC将双链中的mRNA降解,沉默基因转录后的表达。
- 如果miRNA与靶基因mRNA不完全互补,则miRNA将与靶基因mRNA的3'-非翻译区的序列形成非完全互补的杂交双链,miRISC紧紧地结合在杂交双链上,特异性地抑制基因表达。RNA参与了细胞的生长、分化、衰老、调亡、自噬、迁移、侵袭等多种过程。
2.siRNA
siRNA有内源性和外源性之分,内源性siRNA是由细胞自身产生的。外源性siRNA来源于外源入侵的基因表达的双链RNA,经Dicer切割所产生的具有特定长度(21~23bp)和特定序列的小片段RNA。这些siRNA可以与AGO蛋白结合,并诱导这些mRNA的降解。siRNA还有抑制转录的功能。利用这一机制发展起来的RNA干扰(RNA interference,RNAi)技术是用来研究基因功能的有力工具。
3.piRNA
piRNA是从哺乳动物生殖细胞中分离得到的一类长度约为30nt的小RNA。这类小RNA与PIWI蛋白家族成员结合才能发挥其调控作用,故称为piRNA(piwi interacting RNA)。piRNA主要存在哺乳动物生殖细胞和干细胞中,通过与PIWI蛋白家族成员结合形成piwi复合物来调控基因沉默。
(二)IncRNA
长非编码RNA是一类长度为200~100000个核苷酸的RNA分子。它们不编码任何蛋白质。以前它们被认为是基因组转录过程中的“噪声”而被忽略掉,现在越来越多的证据表明它们是一类具有特殊功能的RNA。
IncRNA由RNA聚合酶Ⅱ转录生成,经剪切加工后,形成具有类似于mRNA的结构。IncRNA有poly(A)尾巴和启动子,但序列中不存在可读框。
IncRNA可以来源于蛋白质编码基因、假基因以及蛋白质编码基因之间的DNA序列。IncRNA定位于细胞核内和细胞质内。IncRNA具有强烈的组织特异性与时空特异性,不同组织之间的IneRNA表达量不同,同一组织或器官在不同生长阶段,IneRNA表达量也不同。
IncRNA的作用机制有以下几种:
- 结合在编码蛋白质的基因上游启动子区,干扰下游基因的表达;
- 抑制RNA聚合酶Ⅱ或者介导染色质重构以及组蛋白修饰,影响下游基因的表达;
- 与编码蛋白质基因的转录本形成互补双链,干扰RNA的剪切,形成不同的剪切形式;
- 与编码蛋白质基因的转录本形成互补双链,在Dicer酶的作用下产生内源性siRNA;
- 与特定蛋白质结合,IncRNA转录本可调节相应蛋白质的活性;
- 作为结构组分与蛋白质形成核酸蛋白质复合体;
- 结合到特定蛋白质上,改变该蛋白质的细胞定位;
- 作为小分子RNA(如miRNA、piRNA)的前体分子。
由此可见,IncRA具有调控的多样性,可从染色质重塑、转录调控及转录后加工等多个层面上实现对基因表达进行调控。长非编码RNA与人类疾病的发生密切相关,现已得知,包括癌症以及退行性神经疾病在内的多种严重危害人类健康的重大疾病都与长链非编码RNA的序列和空间结构的异常、表达水平的异常、与结合蛋白相互作用的异常等密切相关。
(三)环状RNA
2012年,美国科学家在研究人体细胞的基因表达时,首次发现了环形RNA分子。截至目前,人们已经在哺乳动物转录组中发现了数以千计的环状RNA,这似乎表明环状RNA而非线性RNA分子是更普遍的现象。
环状RNA是一类特殊的RNA分子。与传统的线性RNA不同,circRNA分子呈封闭环状结构,没有5'-端和3'-端,因此不受RNA外切酶的影响,表达更稳定,不易降解。已知的circRNA分子或来自外显子,或兼有外显子和内含子的部分。circRNA几乎完全定位于细胞核中。circRNA具有序列的高度保守性,具有一定的组织、时序和疾病特异性。由于circRNA的首尾连接,没有尾巴,因此circRNA容易在传统的分离过程被丢弃掉。这是为什么以前circRNA一直没有被发现的主要原因。
小鼠、人类和斑马鱼的各个组织都可以表达circRNA分子,这些circRNA分子富含miRNA的结合位点,在细胞中起到miRNA海绵(miRNA sponge)的作用,通过结合miRNA,进而解除miRNA对其靶基因的抑制作用,升高靶基因的表达水平,产生相应的生物学效应,这一作用机制被称为竟争性内源RNA(competing endogenous RNA,ceRNA)机制。
通过与疾病关联的miRNA相互作用,circRNA在疾病中发挥着重要的调控作用。这说明cireRNA很可能就是一类新的调控性内源竞争性RNA,从而使得环状RNA在作为新型临床诊断标记物的开发应用上具有明显优势。